8.2 REGULACIÓN DE LA TRANSCRIPCIÓN EN
ORGANISMOS PROCARIÓTICOS

Cuando las condiciones de crecimiento son tan
pobres que faltan aminoácidos para la síntesis de proteínas, la respuesta
estricta inhibe la síntesis de rRNA y tRNA 10 a 20 veces, y unas 3 veces la de
mRNA, además de reprimir la síntesis de proteínas y algunas rutas anabólicas.
La falta de aminoácidos la detecta relA
que se encuentra en uno de cada 200 ribosomas. Esta proteína cataliza las
reacciones:
ppG (GDP) + ATP --> ppGpp + AMP
pppG (GTP) + ATP --> pppGpp + AMP
Cuyo fin es la síntesis de la señal (los tetra- y
pentafosfato de guanosina) que desencadenará esta respuesta. La señal
desaparece por la acción de spoT.
- CAMBIOS EN LA ESTRUCTURA DE LA RNA-POLIMERASA

- CAMBIOS EN LA ESTRUCTURA DEL DNA
Metilación
La metilación provoca un cambio de estructura
en el apareamiento entre las bases nitrogenadas que puede alterar su
reconocimiento por algunas proteínas. El más conocido es el de la metilasa dam
que reconoce la secuencia GATC y metila la A.
La mayor parte de los genes cuya expresión se ve reprimida por la metilación son genes cuya expresión sólo se necesita durante la replicación (único momento en el que una cadena del DNA está transitoriamente hemimetilado), permaneciendo reprimidos el resto del ciclo celular.
Los genes mioC y dnaA de E. coli implicados en el inicio de la replicación del DNA son de los pocos que se activan con la metilación, puesto que si el DNA está totalmente metilado, es señal de que se pueden expresar para iniciar otro ciclo replicativo.
Superenrollamiento
Para mantener una situación homeostática en la
célula en relación al número de superenrollamientos es necesario mantener con
una regulación contraria los genes topA (codifica la topoisomerasa I) y gyrA
y gyrB (determinan las dos subunidades de la DNA-topoisomerasa II). No
se conoce el mecanismo molecular que controla esta regulación. Sí se sabe que
mutantes en las topoisomerasas disminuyen la tasa general de transcripción.
- CAMBIOS EN LA INTERACCIÓN ENTRE EL DNA Y LA RNA-POLIMERASA
Lo provocan aquellos cambios
que, sin alterar ni la estructura del DNA ni la de la RNA-polimerasa, sí que
afectan la interacción entre ambas. Es necesaria la comparecencia de una
tercera molécula, habitualmente una proteína (aunque a veces puede ser RNA).
Tres mecanismos pueden alterar esta interacción:
1.- Modificación de la interacción
entre RNA polimerasa y el promotor debido a la existencia de una proteína reguladora y un
efector.
2.- Secuencias reguladoras a
distancia.
3.-Modificación de la
terminación: antiterminación.
Operón
lactosa
Fue el primer operón
descubierto y quizá el mejor conocido. Está compuesto por un
gen regulador I y los genes estructurales Z, Y y A. Se activa por la
presencia de un inductor
(natural como lactosa o alolactosa, o sintético como el IPTG), mientras que lacI
actúa como gen represor.
El control que ejercen ambos puede activar o reprimir
la transcripción del operón. Con la 3ª edición de —Lehninger Principios de
Bioquímica— se distribuye una guía para conocer mejor la estructura del
represor LacIp; es aconsejable visitarla. Este operón también
puede sufrir una activación con el gen CRP , pasando a formar parte del regulón de la
represión catabólica. La proteína reconoce el DNA gracias a un dominio hélice-giro-hélice
(HTH por Helix-Turn-Helix) muy
abundante en procariotas. Este activador dimeriza en presencia de cAMP.
La se establece a través de una estructura del tipo
«cremallera de leucinas» que se verá más adelante. Restablecer aspecto inicial.
Su unión al DNA provoca una torsión en el mismo, lo que facilita el reconocimiento del promotor por la RNA-polimerasa. Compruébalo seleccionando en el menú Jmol «Select->Mouse Click Action->Angle» y pincha sobre un extremo del DNA, un punto central y el otro extremo. En la barra inferior de la ventana te devolverá un ángulo en torno a 120°. Todos los promotores reconocidos por CRP carecen de una buena secuencia -35 (y en algunos casos tampoco la -10), por lo que quizá la presencia de CRP pueda sustituir una reacción eficaz entre la RNA-polimerasa y la región -35. CRP interacciona con la subunidad α de la RNA-polimerasa. Además, mientras LacIp esté unido a su operador, el promotor no funcionará aunque tenga unido CRP. Y el promotor que no tenga LacIp unido no funcionará a menos que CRP se una a su operador.
Se utilizan corrientemente una serie de mutaciones del operón lac con interés biotecnológico, que son:
1.- Mutación M15: afecta a los aminoácidos
N-terminales 11 a 41 del gen lacZ. Estos mutantes son inactivos, pero
pueden recuperar su actividad en presencia de péptidos con los 75 primeros aminoácidos
de LacZ (complementación α). Esto lo utilizan los sistemas de clonación que
identifican los transformantes por el color azul/blanco en presencia de X-Gal.
2.- Mutación lacUV5: cambia
dos pb en la región -10 del promotor del operón que le adapta la secuencia al
consenso perfecto de la región -10. El promotor se vuelve mucho más activo que
el tipo silvestre, y es independiente de la activación por CRP. Se emplea en
sistemas de clonación o expresión constitutiva que no hay que inducir con IPTG.
3.- Mutantes lacOc: mutaciones en el operador que lo hace
irreconocible por LacIp, de manera que se expresan constitutivamente, sin
necesidad de inductor.
Operón
triptófano y atenuación
La estructura y
regulación del operón muestra que la molécula correpresora
es el triptófano, que junto a trpR (el gen regulador) reprimen la
expresión del operón. Además, este operón es un claro ejemplo de regulación
utilizando el acoplamiento entre transcripción y traducción que hay en
procariotas. Este fenómeno se denomina atenuación,
cuyo modo de acción se basa en la formación de estructuras en
horquilla implicadas en la terminación prematura del transcrito. La
disponibilidad o no de triptófano (por ende, Trp-tRNATrp) es la que
va a permitir que el operón se transcriba totalmente (ausencia de Trp) o que en
presencia de Trp cualquier transcripción iniciada se aborte por la formación de
una horquilla terminadora intrínseca en la región TrpL.
Regulación
de las fases lítica y lisogénica del fago lambda
El sistema regulador
temprano de este fago es mucho más complejo que los anteriores, ya que
intervienen varios genes. Del fino equilibrio entre los represores cI y Cro va a depender que
se entre en un ciclo lítico o en un ciclo lisogénico. El dominio de unión al
DNA es también de tipo HTH
en ambas proteínas.
A diferencia de lo que ocurría con CRP, Cro no provoca ninguna torsión en el DNA al unirse a él. Compruébalo seleccionando en el menú Chime «Select -> Mouse Click Action ->Angle» y pincha sobre un extremo del DNA, un punto central y el otro extremo. Está claro que la distancia entre los dos dominios HTH que interaccionan con el operador va a determinar el grado de torsión del DNA unido.
Una de las principales
peculiaridades de esta regulación es la existencia de dos promotores solapados
en sentido opuesto (PR y PRM) que comparten las mimas
señales de regulación: los operadores OR1, OR2 y OR3.
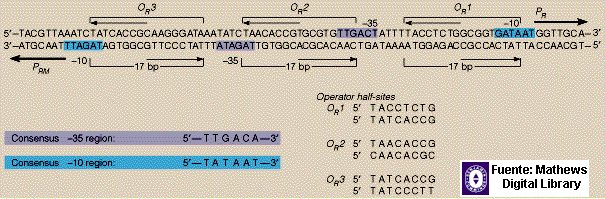
El pasó y/o
mantenimiento de la fase lisogénica depende de que la cantidad de cI
sea suficiente para reprimir los promotores PR y PR sin
afectar la transcripción de él mismo a partir de PRM. El paso a
lisis se debe a la presencia de cro que permitirá la expresión de los
promotores anteriores, pero inhibirá la de PRM.
Regulón
SOS
En una célula sana, lexA
y recA se expresan muy poco, pero lo suficiente como para que LexAp
mantenga reprimidos (inactivados) todos los genes que intervienen en la
respuesta SOS. lexA y recA que son capaces de escapar tenuemente
a esta represión. Cuando hay daño en el DNA, se generan fragmentos de ssDNA
(huecos en dsDNA) que, al ser reconocidos por RecAp, se manifiesta su actividad
proteolítica. De esta manera hidroliza a LexAp, con lo que su concentración
baja, perdiéndose la represión de los genes SOS. Al desaparecer el ssDNA, se
recuperan los niveles basales de LexAp y se reprime el regulón.

Recambio
del mRNA
En procariotas, el mRNA
sufre pocas o ninguna modificación tras su transcripción debido al acoplamiento
con la traducción. Por tanto, lo único que puede considerarse en un recambio
de la molécula. La mayoría de los mRNA tienen una vida media corta (2 a 3
minutos) debido a que estos organismos tienen que adaptarse rápidamente a los
cambios ambientales. El mRNA empieza a ser digerido por la zona 5’ mediante una
endorribonucleasa
que genera distintos fragmentos de RNA. Estos nuevos fragmentos degradados son
accesibles por la exorribonucleasa
II (o polinucleótido fosforilasa, la enzima que S. Ochoa
descubrió con una actividad inversa a la que realiza in vivo) que degrada el
RNA en dirección 3’->5’. Esta exorribonucleasa se detiene ante estructuras
secundarias, por lo que la presencia de cualquier horquilla detiene la
degradación. Por eso es frecuente encontrar una secuencia repetida palindrómica
(REP)
que forma una horquilla en la zona 3' de los genes procariotas.
Procesamiento
del rRNA
Los rRNA se transcriben como
un RNA de mayor tamaño (pre-rRNA y pre-tRNA) que sufren una serie de
procesamientos endonucleolíticos que generarán los RNA maduros. En el genoma de
E. coli hay 7 operones distintos para codificar los rRNA, que se
denominan rrnA-G.
Cada uno contiene en un único transcrito las secuencias de los rRNA 16S, 5S y
23S separados a veces por uno o varios tRNA. La endorribonucleasa III se encarga de
generar p16 y p23, los precursores del 16S y 23S respectivamente. En presencia
de varias proteínas del ribosoma se termina su maduración. Los rRNA sufren metilación, formando
N6,N6-dimetil-adenosina, y O2’-metil-ribosa. La metilación de la ribosa está
relacionada con la protección de los enlaces fosfodíester para evitar la
degradación de los rRNA. Una vez que el rRNA p23 está en el ribosoma se produce
el corte que genera los rRNA maduros 5S y 23S.

Procesamiento
de los tRNA
La mayoría de los tRNA de E.
coli se transcriben en pre-tRNA que contienen de 1 a 7 tRNA cada uno,
separados por secuencias flanqueantes largas. En el extremo 3’ del operón suele
haber un gen traducible. El tRNA de la Tyr es
uno de los mejor conocidos. Se sabe que en la etapa 4 (y antes de que se
modifiquen las bases del tRNA) se puede reconstruir la secuencia CCA en los
casos en los que la exorribonucleasa
D haya degradado demasiado mediante la enzima
tRNA-nucleotidil-transferasa. En algunos tRNA de algunas bacterias (incluida E.
coli) ya se ha descrito el ayuste
(splicing) necesario para eliminar intrones de la secuencia y producir el tRNA
maduro final.
Técnicas
experimentales
El retraso en gel
es una técnica que permite saber si hay alguna proteína que reconozca una
secuencia de DNA determinada. Si tal proteína existe, el complejo DNA-proteína
migra más lentamente en un gel no desnaturalizante, mientras que si el DNA no
es reconocido por ninguna proteína, migra más rápidamente con el frente
electroforético. En la imagen ejemplo se muestra un DNA que es reconocido por
dos proteínas distintas.
No hay comentarios:
Publicar un comentario